

Hieff Canace® Gold High-Fidelity DNA Polymerase基于Pyrococcus Furiosis DNA Polymerase,经基因工程改造而成。该酶具有5’→3’DNA聚合酶活性和3’→5’核酸外切酶活性,其保真性是Taq DNA聚合酶的52倍,是普通Pfu DNA聚合酶的6倍。酶溶液中添加了热启动因子,极大提高了扩增的检出率和产物的特异性。酶溶液中添加了延伸因子使得该酶具有长片段扩增能力,扩增速度达到15 sec/kb,扩增目的片段的长度可长达10 kb。本产品配备了优化缓冲液,添加了PCR增强组分,使得该酶适用于复杂模板的扩增。扩增产物为平末端。
产品组分
| 组分编号 | 组分名称 | 10148ES10 (10U) | 10148ES60 (100U) | 10148ES76 (500U) | 10148ES80 (1,000U) |
|---|---|---|---|---|---|
| 10148-A | Hieff Canace® Gold High-Fidelity DNA Polymerase (2 U/μL) | 5 μL | 50 μL | 250 μL | 250 μL×2 |
| 10148-B | 2×Canace® Gold PCR buffer(含Mg2+,dNTPs) | 300 μL | 1 mL×3 | 1 mL×15 | 1 mL×30 |
- 保真性高:Taq酶的52倍,Pfu酶的6倍。
- 特异性好:目的条带单一。
- 延伸速度快:可达15 sec/kb,极限速度1 sec/kb。
- 灵敏度高:可扩增低至1 pg模板。
- 适用性广:适合不同长度,不同GC含量模板的扩增。
- 高特异性:目的条带单一

图. 5种品牌的高保真DNA聚合酶扩增2 kb,4 kb的小鼠gDNA。Hieff Canace® Gold高保真DNA聚合酶与同类产品对比,显示出更高的特异性和产量。Y:Hieff Canace® Gold高保真DNA聚合酶;A-E:其他品牌的高保真DNA聚合酶;M:1kb DNA Marker。
- 延伸速度快:可达15 sec/kb,极限速度达1 sec/kb
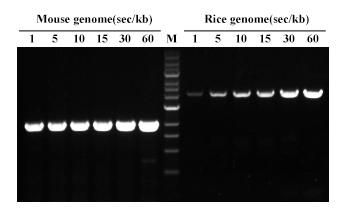
图. Hieff Canace® Gold高保真DNA聚合酶分别扩增小鼠gDNA 2 kb和水稻gDNA 3 kb片段。不同延伸速度(1-60 sec/kb)都可有效扩增。M:1kb DNA Marker。
- 灵敏度高:可扩增低至1 pg模板
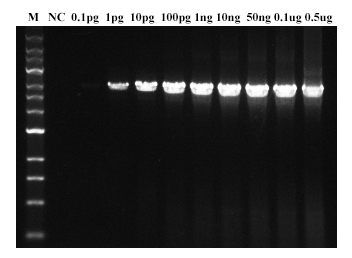
图. Hieff Canace® Gold高保真DNA聚合酶扩增质粒3 kb片段,不同模板量(1 pg-0.5 μg)都可有效扩增。NC:阴性对照。M:1kb DNA Marker。
- 适用于不同GC含量片段的扩增
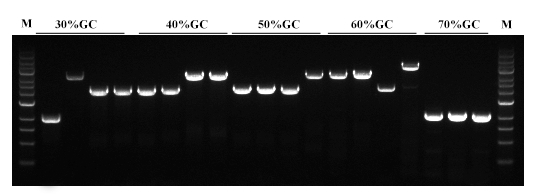
图. Hieff Canace® Gold高保真DNA聚合酶对不同GC含量(30-70%)片段都可高效率的进行扩增。M:1kb DNA Marker。
- 适用于不同长度片段的扩增
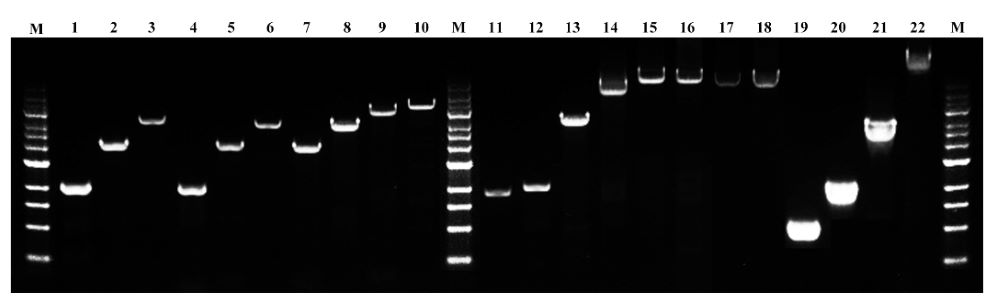
图. Hieff Canace® Gold高保真DNA聚合酶对不同模板、不同长度(1-12 kb)片段都可高效率的进行扩增。M:1kb DNA Marker。模板:1-3:拟南芥gDNA,4-6:水稻gDNA,7-9:小鼠gDNA,10:人cDNA,11-18:人gDNA,19-21:质粒DNA,22:λDNA。
冰袋运输。-20℃保存,有效期2年。
1. hu M, Dai X. Growth suppression by altered (p) ppGpp levels results from non-optimal resource allocation in Escherichia coli[J]. Nucleic acids research, 2019, 47(9): 4684-4693. (IF11.6)
2. Guo L, Yang W, Huang Q, et al. Selenocysteine-specific mass spectrometry reveals tissue-distinct selenoproteomes and candidate selenoproteins[J]. Cell chemical biology, 2018, 25(11): 1380-1388. e4.(IF6.762)
3. Li Y, Xu W, Zhang F, et al. The Gut Microbiota-Produced Indole-3-Propionic Acid Confers the Antihyperlipidemic Effect of Mulberry-Derived 1-Deoxynojirimycin[J]. Msystems, 2020, 5(5).(IF6.633)
4. He H, Wu S, Mei M, et al. A combinational strategy for effective heterologous production of functional human lysozyme in Pichia pastoris[J]. Frontiers in Bioengineering and Biotechnology, 2020, 8. (IF3.644)
5. He H, Zhai C, Mei M, et al. Functional expression of porcine interferon-α using a combinational strategy in Pichia pastoris GS115[J]. Enzyme and microbial technology, 2019, 122: 55-63. (IF3.448)
6. Liu Z, Lv X, Song E, et al. Fostered Nrf2 expression antagonizes iron overload and glutathione depletion to promote resistance of neuron-like cells to ferroptosis[J]. Toxicology and Applied Pharmacology, 2020, 407: 115241. (IF3.347)
7. Liu Y, Shen Q, Zhao X, et al. Cell-free mitochondrial DNA in human follicular fluid: a promising bio-marker of blastocyst developmental potential in women undergoing assisted reproductive technology[J]. Reproductive Biology and Endocrinology, 2019, 17(1): 54.(IF3.235)
8. He L, Li J, Peng P, et al. Genomic analysis of a Chinese MDV strain derived from vaccine strain CVI988 through recombination[J]. Infection, Genetics and Evolution, 2020, 78: 104045.(IF2.611)



